

Fourth Ventricle

Abstract
OBJECT The two most common surgical routes to the fourth ventricle are the transvermian and telovelar approaches. The purpose of this study was to compare the microanatomy and exposures gained through these approaches.
METHODS Ten formalin-fixed specimens were dissected in a stepwise manner to simulate the transvermian and telovelar surgical approaches. Stealth image guidance was used to compare the exposures and working angles obtained using these approaches.
The transvermian and telovelar approaches provided access to the entire rostrocaudal length of the fourth ventricle floor from the aqueduct to the obex. In addition, both approaches provided access to the entire width of the floor of the fourth ventricle. The major difference between the two approaches regarded the exposure of the lateral recess and the foramen of Luschka. The telovelar, but not the transvermian, approach exposed the lateral and superolateral recesses and the foramen of Luschka. The transvermian approach, which offered an incision through at least the lower third of the vermis, afforded a modest increase in the operator’s working angle compared with the telovelar approach when accessing the rostral half of the fourth ventricle.
CONCLUSIONS The transvermian approach provides slightly better visualization of the medial part of the superior half of the roof of the fourth ventricle. The telovelar approach, which lacks incision of any part of the cerebellum, provides an additional exposure to the lateral recesses and the foramen of Luschka.

ATLAS Choice Bipolar Forceps
Designed for your every surgical maneuver
Five tip sizes for brain and spine procedures
Unparalleled non-stick and low-profile features
Introduction
The two most common surgical approaches to the fourth ventricle are the transvermian and telovelar approaches. The first involves incising the inferior vermis of the cerebellum and retracting the two halves of the vermis in opposite lateral directions.1,3,6,11,12 In the second approach, the tela choroidea and inferior medullary velum, which form the lower half of the roof of the fourth ventricle, are opened and the lower vermis is retracted as a unit to provide exposure into the fourth ventricle.17,20,22 The technical differences between the two approaches and the anatomical structures that limit the surgical view in each approach result in differences with regard to the exposure of the fourth ventricle floor, the lateral recess, and the foramen of Luschka. The purpose of this study was to describe and compare the microanatomical features of the transvermian and telovelar approaches and to relate these features to the exposure gained through each approach.
Materials and Methods
Ten formalin-fixed specimens, in which the arteries had been perfused with red silicone and the veins with blue silicone, were examined in this study. The specimens were dissected in a stepwise manner, with the aid of an operating microscope set to 3 to 40, to simulate exposures that can be obtained using the transvermian and telovelar approaches. Special attention was directed to the anatomical structures that limit visualization in each approach.
Three of 10 cadaveric heads were studied using MR imaging, and the data were registered on a Stealth workstation by using standard protocols. Magnetic resonance images were obtained using a 3-tesla MR imaging system (Magnetom Allegra; Siemens Medical Solutions, Malvern, PA) and transferred to a Stealth Image Guidance workstation (Medtronic Surgical Navigation Technologies, Louisville, CO). Ten fiducial markers used for image registration were attached to the skin overlying the superior sagittal sinus and the parietooccipital area. The head was placed similar to the orientation provided by the semisitting position and a suboccipital craniotomy was performed through a midline vertical skin incision. A 3D model was built and a dynamic reference array was attached to the cadaver. The specimen was registered to the 3D model and image, resulting in an estimated accuracy of better than 2 mm. An active probe was used to indicate the surgical exposure on the 3D model and on three orthogonal MR images. These images were recorded through screen captures. The superior, inferior, lateral, superolateral, and foramen of Luschka exposures through each approach to the fourth ventricle were determined using image guidance under direct microscopic visualization.
We compared the operative angle of approach through the transvermian and telovelar routes. The angle was measured from the floor of the fourth ventricle in the sagittal plane. The apex of the angle was placed at three sites along the ventricle floor: the aqueduct, the midpoint (50% point) of the distance measured from the obex to the inferior border of the aqueduct, and the point 75% of the same distance.
Results
Anatomical Relationships
Suboccipital Surface
Operative approaches to the fourth ventricle move through the suboccipital or posterior surface of the cerebellum, which is located below the transverse sinus and between the sigmoid sinuses. The vermis is folded into and forms the suboccipital cortical surface within the posterior cerebellar incisura, which is a deep vertical depression on the posterior surface of the cerebellum (Fig. 1). The vermian surface within the posterior cerebellar incisura has a shape similar to that of a diamond, the upper portion of which is called the pyramid. The lower half of the vermian surface projects downward between the tonsils and is called the uvula, thus mimicking the anatomy found in the oropharynx (Fig. 1). The medial margin of the rostral portion of the tonsils borders the tapering edges of the uvula. The strip of vermis within the incisura is broadest at the junction of the pyramid and uvula. Inferiorly, the posterior cerebellar incisura is continuous with the vallecula, a cleft between the tonsils that opens into the foramen of Magendie.
The tonsils, the most prominent structures blocking access to the caudal portion of the fourth ventricle, are considered hemispherical components. The tonsils are ovoid structures, which, along their superolateral borders, are attached to the remainder of the cerebellar hemisphere, whereas the remaining parts of each tonsil are free surfaces (Fig. 1). The anterior and medial surfaces of each tonsil are important landmarks that are recognizable when approaching the fourth ventricle. The anterior surface faces the cerebellomedullary fissure, which is a cleft between the tonsil and the posterior surface of the medulla oblongata. The medial surfaces of the tonsils face each other across a narrow cleft, the vallecula, which leads into the inferior portion of the floor of the fourth ventricle.
Roof
The roof of the fourth ventricle is a tent-shaped structure. The apex of the roof, the fastigium, is the site of the roof’s greatest height and width. The fastigium divides the roof into superior and inferior portions. The superior half of the roof expands laterally and posteriorly from the aqueduct to the level of the fastigium and lateral recess (Figs. 2 and 3). The superior half of the roof is formed medially by the superior medullary velum, which connects at the fastigium with the inferior medullary velum and laterally with the inner surfaces of the superior and inferior cerebellar peduncles. The fibers of the superior cerebellar peduncle arise in the dentate nucleus and ascend to form the ventricular surface of the superior portion of the lateral wall (Fig. 3).
The inferior portion of the roof is formed by the tela choroidea and the inferior medullary velum (Figs. 2 and 4). The inferior medullary velum is a membranous remnant of the connection between the nodule and the folliculi that forms the flocculonodular lobe.9 It is a thin bilateral layer of neural tissue that extends from the nodule medially and blends into the dorsal margin of each lateral recess, forming the peduncle of each flocculus laterally (Figs. 2 and 4). Caudally, the inferior medullary velum attaches to the tela choroidea. The tela choroidea contains a vascular layer of choroidal arteries and veins between its layers and the choroid plexus projects from its ventricular surface (Fig. 4). Importantly, no known functional neural tissue resides within the structures that form the inferior portion of the roof of the fourth ventricle.
The telovelar junction, the line of attachment of the inferior medullary velum to the tela choroidea, extends laterally from the nodule into each lateral recess (Fig. 4). The tela attaches to the inferolateral edges of the floor along narrow ridges, the taeniae, which meet at the obex. The tela choroidea has three openings to the subarachnoid recess: the paired foramina of Luschka, located at the lateral recess, and the foramen of Magendie, located medially at the caudal end of the fourth ventricle (Fig. 4).

FIG. 1. Photographs demonstrating posterior views of cadaveric brain specimens. The suboccipital surface of the cerebellum and the cerebellomedullary fissure are shown. Panels A and B show one specimen and C and D another. A: The suboccipital surface is located below and between the sigmoid and transverse sinuses and is the surface exposed during a wide bilateral suboccipital craniotomy. The vermis sits in a depression, the posterior cerebellar incisura that lies between the hemispherical surfaces. The cerebellomedullary fissure extends superiorly between the cerebellum and the medulla oblongata. The vallecula extends upward between the tonsils and communicates through the foramen of Magendie with the fourth ventricle. The left PICA courses below the lower pole of the tonsil to reach the lower margin of the cerebellomedullary fissure and supplies part of the suboccipital surface. B: The left tonsil has been retracted laterally and the uvula medially to expose the inferior portion of the roof of the fourth ventricle. The inferior portion of the roof is formed by the tela choroidea and the inferior medullary velum. The bifurcation of the PICA into the medial and lateral trunks has been retracted to show the lateral recess. Some branches of the medial trunk, which supplies the vermis, course along the uvulotonsillar space along with the vein of the cerebellomedullary fissure. C: The right tonsil has been retracted medially from the biventral lobule. The tonsils, the most prominent structures blocking access to the caudal portion of the fourth ventricle, are attached to the cerebellar hemisphere along their superolateral borders. The fissure between the tonsil and the biventral lobule is called the tonsillobiventral fissure. D: Enlarged posterior view of the same hemisphere. Both tonsils have been retracted laterally to expose the inferior medullary velum and the tela choroidea. The choroidal arteries course along the tela choroidea, from which the choroid plexus projects into the roof of the fourth ventricle. The inferior vermian surface within the posterior cerebellar incisura has a diamond shape, the upper portion of which is called the pyramid. The lower half projects downward between the tonsils and is called the uvula. The nodule is hidden deep in relation to the uvula. The medial margin of the rostral portion of the tonsils borders the tapering edges of the uvula. The strip of the vermis within the incisura is broadest at the junction of the pyramid and uvula. The right PICA bifurcates into lateral and medial trunks, which course along the uvulotonsillar space. The lateral recess, a narrow pouch formed by the union of the roof and floor, is covered caudally by the tela choroidea. A. = artery or arteries; Cer. = cerebellar or cerebello; Chor. = choroidal, Fiss. = fissure; For. = foramen; Inf. = inferior; Lat. = lateral; Med. = medial or medullary; Post. = posterior; Tons. Bivent. = tonsillobiventral; Tr. = trunk.
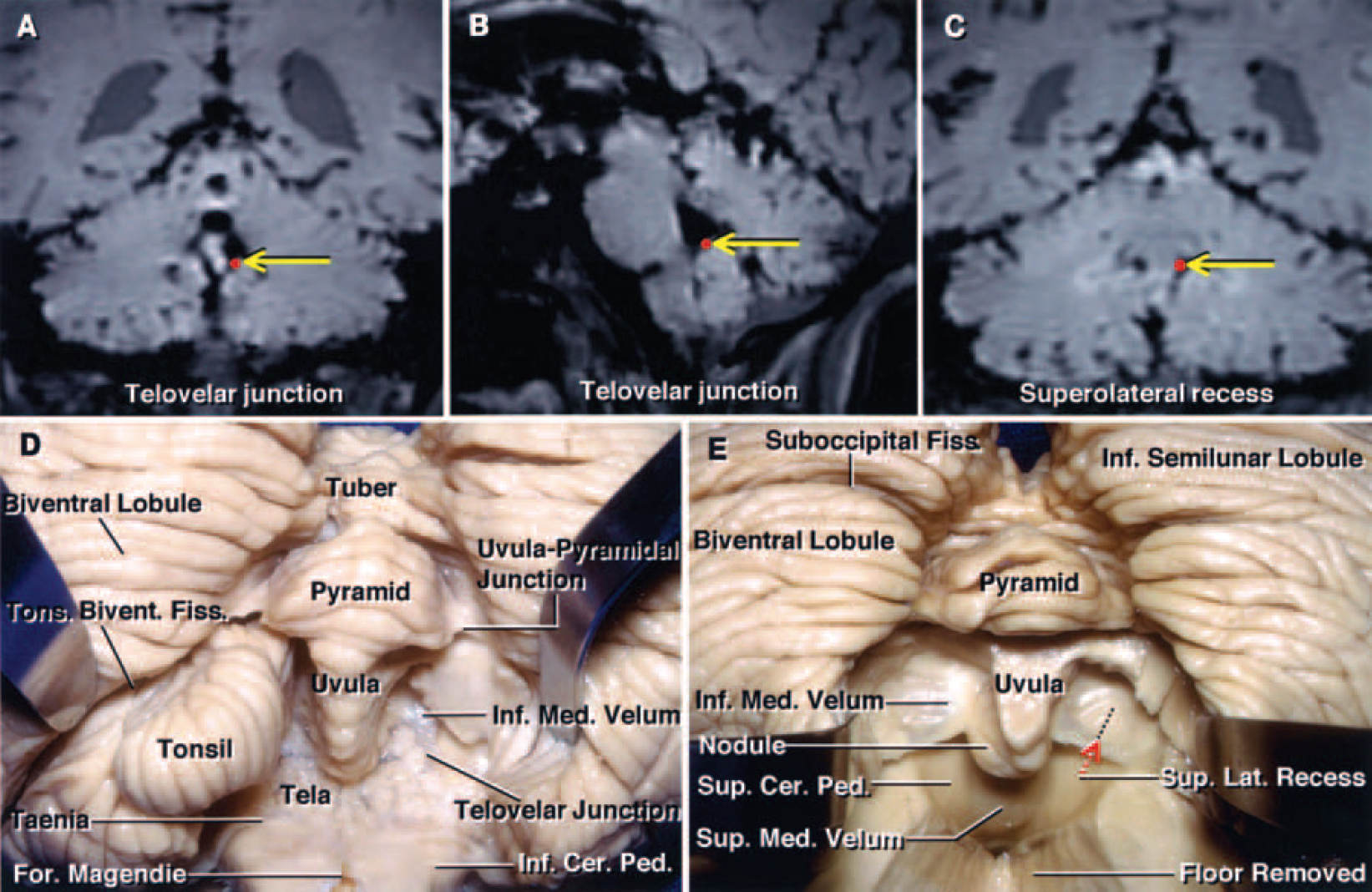
FIG. 2. A–C: Coronal (A and C) and sagittal (B) MR images of a cadaveric brain obtained from the Stealth workstation, demonstrating the location of the telovelar junction (red dots indicated by yellow arrows in A and B) and the superolateral recess (red dot indicated by yellow arrow in C). D and E: Photographs of cadaveric brain specimens showing the stepwise dissection of the suboccipital surface of the cerebellum and the inferior portion of the fourth ventricle. D: The left biventral lobule has been retracted to show the tonsillobiventral fissure. The right tonsil has been removed by dividing the peduncle of the tonsil located along the superolateral margin of the tonsil. The uvula projects downward between the tonsils, mimicking the anatomy in the oropharynx. The uvulopyramidal junction, the broadest strip of vermis within the posterior cerebellar incisura, limits the lateral retraction of the tonsils. The telovelar junction is the line of attachment of the tela to the velum. E: Both tonsils and part of the floor have been removed to expose the superior medullary velum and the superolateral recess. The inferior medullary velum is a thin bilateral layer of neural tissue that extends from the nodule medially and blends into the dorsal margin of each lateral recess, forming the peduncle of each flocculus laterally. The incision through the inferior portion of the vermis during the transvermian approach exposes the underlying nodule, which must be incised to gain access to the fourth ventricle. This incision through the vermis is limited by the position of the superior medullary velum, which is closely related to the anterior portion of the nodule. The superior medullary velum forms the medial part of the superior half of the roof and expands laterally and posteriorly from the aqueduct to the level of the fastigium and lateral recess. The superolateral recess, located within the fourth ventricle and lateral to the uvula and nodule, is medial to the superior cerebellar peduncles and lies anterior to the upper edge of the inferior medullary velum. Ped. = peduncle; Sup. = superior.
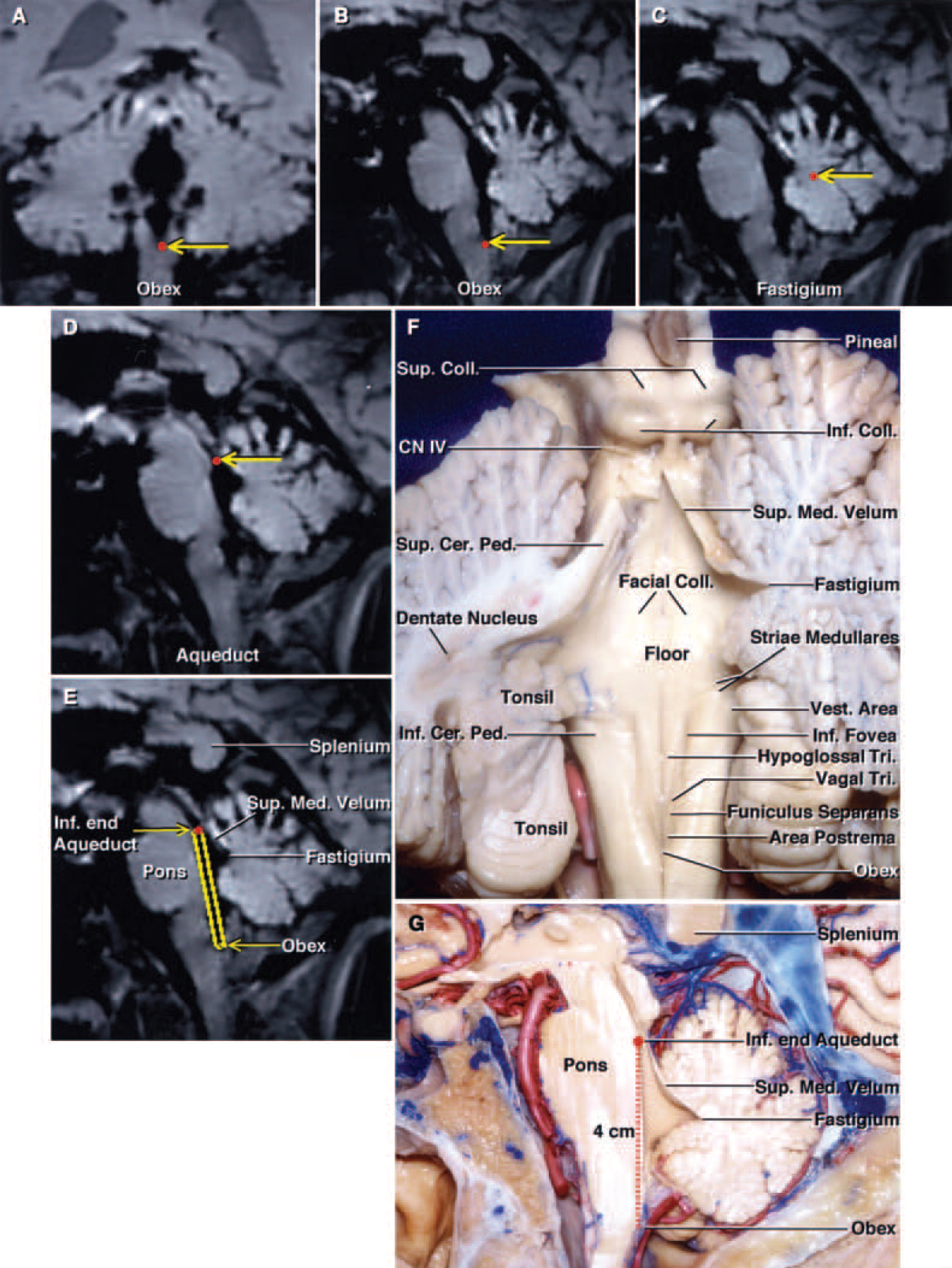
FIG. 3. A–E: Stealth MR images demonstrating the anatomical landmarks (red dots indicated by yellow arrows) along the floor and roof of the fourth ventricle. Coronal (A) and sagittal MR images (B) demonstrating the location of the obex. Sagittal MR images (C and D) revealing the fastigium and aqueduct. Sagittal MR image (E) depicting the floor of the fourth ventricle and its relation to the superior medullary velum and fastigium. (This brain section is located at approximately the same level as the sagittal section of a cadaver shown in panel G.) F: The cerebellum has been cut at the midline and the left half has been sectioned further in an oblique coronal plane to show the relationship of the rostral pole of the tonsil to the dentate nucleus. The dentate nucleus appears to wrap around the tonsil. The fibers of the superior cerebellar peduncle arise in the dentate nucleus and ascend in the medial side of the superior cerebellar peduncle. The striae medullaris cross the midportion of the floor. The vestibular area, the portion of the floor that is lateral to the median eminence and the sulcus limitans, is located at the lateral limit of the floor of the fourth ventricle. The inferior fovea is a depression in the sulcus limitans located lateral to the hypoglossal triangle. The median eminence contains the facial colliculi in its upper part and the hypoglossal and vagal triangles and the area postrema in its lower portion. The median eminence is crossed by the funiculus seperans. The trochlear nerves arise below the inferior colliculus. G: Sagittal section of a hemisphere. The full length of the floor of the fourth ventricle from the inferior edge of the aqueduct to the obex is approximately 4 cm. The roof of the fourth ventricle is a tent-shaped structure. The apex of the roof, the fastigium, is the site of the roof’s greatest height and width. The superior half of the roof expands posteriorly from the aqueduct to the level of the fastigium. This sagittal section corresponds to the Stealth MR image in panel E. CN = cranial nerve; Coll. = colliculi or colliculus; Tri. = triangle; Vest. = vestibular.

FIG. 4. A–C: Coronal (A), sagittal (B), and axial (C) MR images of a cadaveric head obtained from the Stealth workstation, demonstrating the lateral recess (red dots indicated by yellow arrows). The point marked on all three images is identical and was obtained from the same cadaver. D and E: Photographs of another cadaveric specimen showning the stepwise dissection of the suboccipital surface of the cerebellum. Both tonsils have been retracted superolaterally and the left half of the tela choroidea has been opened (D). The choroid plexus projects from the ventricular surface of the tela choroidea into the fourth ventricle. The tela choroidea extends laterally over the inferior cerebellar peduncles and forms the posterior wall of the lateral recesses. View of structures shown in panel D (E). The tela choroidea has been retracted to the right side to expose the entire floor of the fourth ventricle and the aqueduct. The fourth ventricle is continuous with the cerebellopontine angle through the foramen of Luschka at the lateral recess. The superior half of the roof is formed medially by the superior medullary velum and laterally by the inner surfaces of the superior cerebellar peduncles. The inferior medullary velum blends into the peduncles of the flocculus laterally and the surface of the nodule medially. F: The left cerebellar hemisphere, the vermis, and the medial portion of the right cerebellar hemisphere have been removed to expose the full length of the floor of the fourth ventricle. The trochlear nerves arise inferior to the inferior colliculus. The superior and inferior cerebellar peduncles face the surface of the ventricle. The median sulcus divides the floor vertically in the midline. The sulcus limitans divides each half of the floor of the fourth ventricle longitudinally into a medial strip, the median eminence, and a lateral portion, the vestibular area. The sulcus limitans is most prominent at the level of two dimples, the superior and inferior fovea. The facial colliculus is medial to the superior fovea and median eminence. The locus ceruleus, a darkened area of cells, is located at the rostral end of the sulcus limitans. G: Anterolateral view of the brainstem and petrosal surface of the cerebellum, corresponding to the area demonstrated by the Stealth MR image in panel A. The choroid plexus protrudes from the lateral recess and foramen of Luschka behind the glossopharyngeal and vagus nerves. The trigeminal nerve arises from the midpons. The abducent nerve arises in the medial portion and the facial and vestibulocochlear nerves at the lateral end of the pontomedullary sulcus. The hypoglossal nerves arise anterior to the olives and the glossopharyngeal, vagus, and accessory nerves arise posterior to the olives. The anterior inferior cerebellar artery arises at the pontine level and courses by the abducent, facial, and vestibulocochlear nerves. The right PICA arises at the level of the vertebrobasilar junction, courses in relation to the lower cranial nerves, and passes dorsally around the medulla oblongata near the lower portion of the tonsil. H: Enlarged view of the foramen of Luschka with the glossopharyngeal and vagus nerves retracted to expose the foramen and the structures forming the lateral recess. The flocculus protrudes laterally from the inferior margin of the middle cerebellar peduncle dorsal to the facial and vestibulocochlear nerves. The rhomboid lip, a sheet of neural tissue attached to the lateral margin of the floor of the fourth ventricle, has been retracted to show the choroid plexus protruding from the lateral recess and the foramen of Luschka. A.I.C.A. = anterior inferior cerebellar artery; Bas. = basilar; Mid = middle; Plex. = plexus; Sul. = sulcus; V. = vein; Vert. = vertebral.
Cerebellomedullary Fissure and Its Vascular Relations
The inferior half of the roof of the fourth ventricle is intimately related to the cerebellomedullary fissure. The fissure has ventral and dorsal walls. The ventral wall is formed by the posterior surface of the medulla, the inferior medullary velum, and the tela choroidea (Fig. 1). The dorsal wall is formed by the uvula, tonsils, and biventral lobules. The cerebellomedullary fissure extends superiorly to the level of the lateral recess and is continuous with the fourth ventricle through the foramen of Magendie and the cisterna magna around the superior surface of the tonsils; the fissure communicates with the cerebellopontine fissure by way of the foramina of Luschka (Fig. 4).
The PICA is intimately related to the cerebellomedullary fissure, the inferior half of the roof of the fourth ventricle, and the suboccipital surface. The PICA arises from the vertebral artery at the anterolateral margin of the medulla oblongata, close to the lower cranial nerves, and courses through a series of deep fissures that lie among the tonsil, vermis, and cerebellar hemisphere (Fig. 1). The PICA is divided into five segments: the anterior medullary, lateral medullary, tonsillomedullary, telovelotonsillar, and cortical segments.15 The PICA segment that is most commonly exposed during the transvermian and telovelar approaches is the telovelotonsillar segment, which courses in the cleft that lies between the tonsil (below) and the tela choroidea and velum (above). The PICAs bifurcate into medial and lateral trunks in their passage around the tonsil (Fig. 1). The medial trunk supplies the vermis and the lateral trunk supplies most of the hemispherical and tonsillar surfaces. Both trunks are frequently encountered during approaches to the fourth ventricle.
The vein of the cerebellomedullary fissure is the largest vein crossing the inferior portion of the fourth ventricle. It originates on the lateral edge of the nodule and uvula, courses laterally along the telovelar junction, and passes caudal to the cerebellar peduncles to reach the cerebellopontine angle. It drains into veins that empty into the superior petrosal sinus.18 The tributaries of the vein of the cerebellomedullary fissure drain portions of the tonsil, inferior vermis, inferior medullary velum, tela choroidea, and attached choroid plexus, as well as the ventricular white matter and dentate nuclei.
Floor and Lateral Recesses
The floor of the fourth ventricle, a rhomboid structure is situated with its rostral two thirds posterior to the pons and its caudal one third posterior to the medulla oblongata (Figs. 2 and 4). Its cranial apex is at the level of the cerebral aqueduct, and caudally it ends at the obex, which lies anterior and deep with respect to the foramen of Magendie. The intermediate portion of the floor of the fourth ventricle extends into the lateral recesses. The vestibular area, the portion of the floor lateral to the median eminence and sulcus limitans, is located at the lateral limit of the floor of the fourth ventricle (Fig. 3). The vestibular nuclei lie beneath the vestibular area, which forms a rounded elevation that extends directly into the lateral recess. Rostral to the vestibular area, the striae medullaris course transversely from the region of the lateral recess across the inferior cerebellar peduncle toward the midline (Fig. 3).
The fourth ventricle is continuous with the cerebellopontine angle through the foramen of Luschka at the lateral recess, which is a narrow pouch formed by the union of the roof and the floor (Fig. 4). The ventral wall of each lateral recess is formed by the junctional portion of the floor and the rhomboid lip, a sheetlike layer of neural tissue. The rostral part is formed by the caudal margin of the cerebellar peduncles. The caudal part of each lateral recess is formed by the tela choroidea, which extends from the lateral part of the taenia to the flocculus (Fig. 4).
Transvermian Approach
The oldest and most widely used surgical approach to the fourth ventricle consists of splitting the inferior vermis on the suboccipital surface.1,11,12 The extent of the incision through the inferior vermis has been poorly described. The incision extends a variable distance through the uvula, pyramid, tuber, and folium of the vermis, depending on the location and size of the pathological area. Most authors advocate limiting the vermian incision to the smallest possible length necessary to gain access to avoid complications associated with splitting the vermis.23 Similarly, a transvermian incision through the inferior vermis and extending into the fourth ventricle was performed in our study. We maximized the transvermian incision to include the inferior vermis up to the fastigium, but did not extend it into the inferior edge of the superior medullary velum. Functionally, the decussating fibers of the superior cerebellar peduncle lie deep in relation to the superior medullary velum, which is a thin lamina of white substance between the superior cerebellar peduncles. The vermian incision exposed the underlying nodule, which was necessarily incised along with the tela choroidea and inferior medullary velum to gain access to the fourth ventricle (Fig. 5).
Retraction of the two halves of the lower vermis provided approximately 1 to 2 cm of working space between the two edges of the inferior vermis. Following the retraction of the vermis and tonsils laterally, it was possible to follow the course of the telovelotonsillar segments of the bilateral PICA and work in between the right and left segments without sacrificing any of the major branches of the vessels (Fig. 5). The vein of the cerebellomedullary fissure coursed horizontally at the site of the midline vermian incision along the tela choroidea and inferior medullary velum in three hemispheres (Fig. 5). After the the tela choroidea and inferior medullary velum had been opened through a midline incision, the full length of the floor (mean length 4 cm) from the aqueduct to obex, could be exposed (Fig. 5). Laterally, the limit of exposure was maximal at the level of the striae medullaris. The initial medial portion of the vestibular area was the lateral limit of exposure. Superiorly, the middle portion of the superior medullary velum was the limit of exposure. At this point in the dissection, exposure of the lateral recess and the foramen of Luschka was not possible in any of our specimens by using the transvermian approach. Exposure of the lateral recess requires removal of a portion of the vermis, whereas exposure of the foramen of Luschka requires additional retraction of the tonsillar pole.3,13
Stealth image guidance was introduced into the dissection field after we obtained maximal exposure through the fourth ventricle (Fig. 5). The mean operative angles of the transvermian approach measured at three locations along the floor of the fourth ventricle were 38 ̊ at the aqueduct, 84 ̊ at the midpoint of the distance from the obex to the aqueduct, and 72 ̊ at the 75% point. The superior medullary velum was the main anatomical structure limiting the working angle in the sagittal plane. The limitation was most significant when we exposed the rostral one fourth of the fourth ventricle, and the operative view decreased when we moved in the mediolateral direction. The fastigium and the inferior portion of the superior medullary velum limited the working angle at the point 75% of the distance between the obex and aqueduct. At the midpoint along the length of the floor of the fourth ventricle, the working angle increased dramatically. At this point, the exposure was limited superficially at the level of the superior limit of the vermian incision.

FIG. 5. A–D: Photographs of cadaveric specimens showing the transvermian approach to the fourth ventricle. The suboccipital surface of the cerebellum (A). The oldest and most widely used surgical approach to the fourth ventricle consists of splitting the inferior vermis on the suboccipital surface. Enlarged view of some of the anatomical structures shown in panel A (B). Both tonsils have been retracted laterally to expose the inferior portion of the vermis, the pyramid, and the uvula. The incision used for the transvermian approach into the fourth ventricle is most commonly described as being through the inferior portion of the vermis (dotted line). Both PICAs course around the lower pole of the tonsil to reach the lower margin of the cerebellomedullary fissure. The inferior portion of the vermis has been incised, and the vermis and tonsils have been retracted laterally (C). The incision through the vermis exposes the underlying nodule, which must be incised to expose the inferior portion of the roof. Retraction of the two halves of the lower vermis provides approximately 1 to 2 cm of working space between the two retracted edges of the inferior vermis. Following the retraction of the vermis and tonsils laterally, it is possible to follow and work between the right and left telovelotonsillar segments of the bilateral PICAs, without sacrificing any of the major branches of the vessels. The vein of the cerebellomedullary fissure courses transversely along the inferior medullary velum. Retraction of the two halves of the nodule and the opening of the tela choroidea and inferior medullary velum through a midline incision provide access to the full length of the floor (mean length 4 cm) from the aqueduct to the obex (D). The transvermian approach provides a wide exposure of the midline superior half of the roof of the fourth ventricle and the superior medullary velum. E–H: Stealth MR images of the transvermian dissection demonstrating the superior, inferior, and lateral limits of exposure (red dots indicated by yellow arrows). The superior limit of the transvermian exposure is at the inferior edge of the aqueduct (E). The inferior limit of the transvermian exposure is at the obex (F) and the right and left lateral limits are at the level of the striae medullaris (G and H). The medial edge of the vestibular area is the lateral limit of exposure. I and J: Sagittal (I) and coronal (J) Stealth MR images obtained during the dissection, demonstrating the trajectory of the transvermian approach (yellow rectangles leading to red dots). The fastigium and the inferior portion of the superior medullary velum are the main anatomical structures that limit the working angle to the rostral one fourth of the floor of the fourth ventricle during the transvermian approach and this is demonstrated in the sagittal plane.
Telovelar Approach
Following exposure of the suboccipital surface of the cerebellum, the uvulotonsillar and medullotonsillar spaces on both sides were opened along with the lateral and deep dissection around the surfaces of the tonsil and the biventral lobule without dividing any neural tissue (Figs. 6–8). The inferior edge of the tonsils was exposed and the dissection extended between the medial side of the tonsil and the adjacent edge of the uvula. The surface of the tonsil that faced the uvula was retracted superolaterally toward the side of the approach, and the uvula was retracted to the opposite side to expose the inferior medullary velum and tela choroidea. Special attention was directed to the location and bifurcation of the PICA into medial and lateral trunks. The PICA bifurcation was exposed near the inferior end of the uvulotonsillar space, around the telovelar junction, in three of the 10 specimens. If the bifurcation was present along the site of incision, it was easier to retract it laterally than medially before we made the incision along the tela and inferior medullary velum (Figs. 6 and 8).
The telovelar incision can be divided in three parts. The first part of the incision, which opens the tela choroidea, begins inferiorly near the foramen of Magendie in the lower portion of the ventricle roof and extends upward to the level of the junction of the tela with the inferior medullary velum (Fig. 6). After we opened the tela choroidea, the full length of the floor of the fourth ventricle came into view in some hemispheres. The second part involves extending the incision superiorly through the inferior medullary velum, which is exposed in the depths of the uvulotonsillar space. The vein of the cerebellomedullary fissure, which crosses the inferior medullary velum, was transected in some specimens. Incising the inferior medullary velum exposed the ipsilateral superolateral recess and provided access to the entire floor of the fourth ventricle in all hemispheres (Figs. 6 and 8). Opening the inferior medullary velum also increased the exposure of the lower portion of the superior medullary velum near the fastigium. Opening the tela and the inferior medullary velum through the uvulotonsillar space on one side provided limited exposure to the lateral portion of the opposite half of the floor; however, shifting the exposure to the opposite uvulotonsillar space and opening the tela and velum provided an identical exposure to that obtained on the other side. The third incision, which was directed between the tonsil and medulla oblongata through the tela forming the lower posterior wall of the lateral recess, provided additional access to the full length of the lateral recess and the foramen of Luschka (Fig. 8).
Stealth image guidance was used to demonstrate the anatomical exposure gained through the telovelar approach (Figs. 7 and 9). The mean operative angles for the telovelar approach at the three sites along the floor of the fourth ventricle were 30 ̊ at the aqueduct, 67 ̊ at the 75% point, and 84 ̊ at the midpoint. The main limitations of the exposure through this approach were the superior medullary velum and the junction between the pyramid and uvula (Figs. 7–9). The junction of the uvula and pyramid, the broadest strip of vermis within the posterior incisural space, limited the lateral and medial retractions of the tonsil and the uvula at the uvulotonsillar space, respectively (Figs. 3 and 8). The operative angle was limited by the uvulopyramidal junction in all specimens during the telovelar approach.
In both the transvermian and telovelar approaches, the operative angle of approach was limited by the superior medullary velum, which does not appear to be a uniform structure when one moves in a mediolateral direction. The superior medullary velum forms a larger angle with the floor of the fourth ventricle in the midline than it does laterally. In the telovelar approach, the fourth ventricle is approached off the midline through the superolateral recess, which accounts in part for the decrease in operative angle. The mean differences between the operative angles of the two approaches did not exceed 8 ̊ in favor of the transvermian approach.
The transvermian and telovelar approaches provide excellent exposure of the entire floor of the fourth ventricle. The transvermian approach, performed using a midline vermian incision, provided a slightly better exposure of the midline superior half of the roof of the fourth ventricle and fastigium. The telovelar approach provided additional access to the lateral recesses and foramen of Luschka in all specimens.
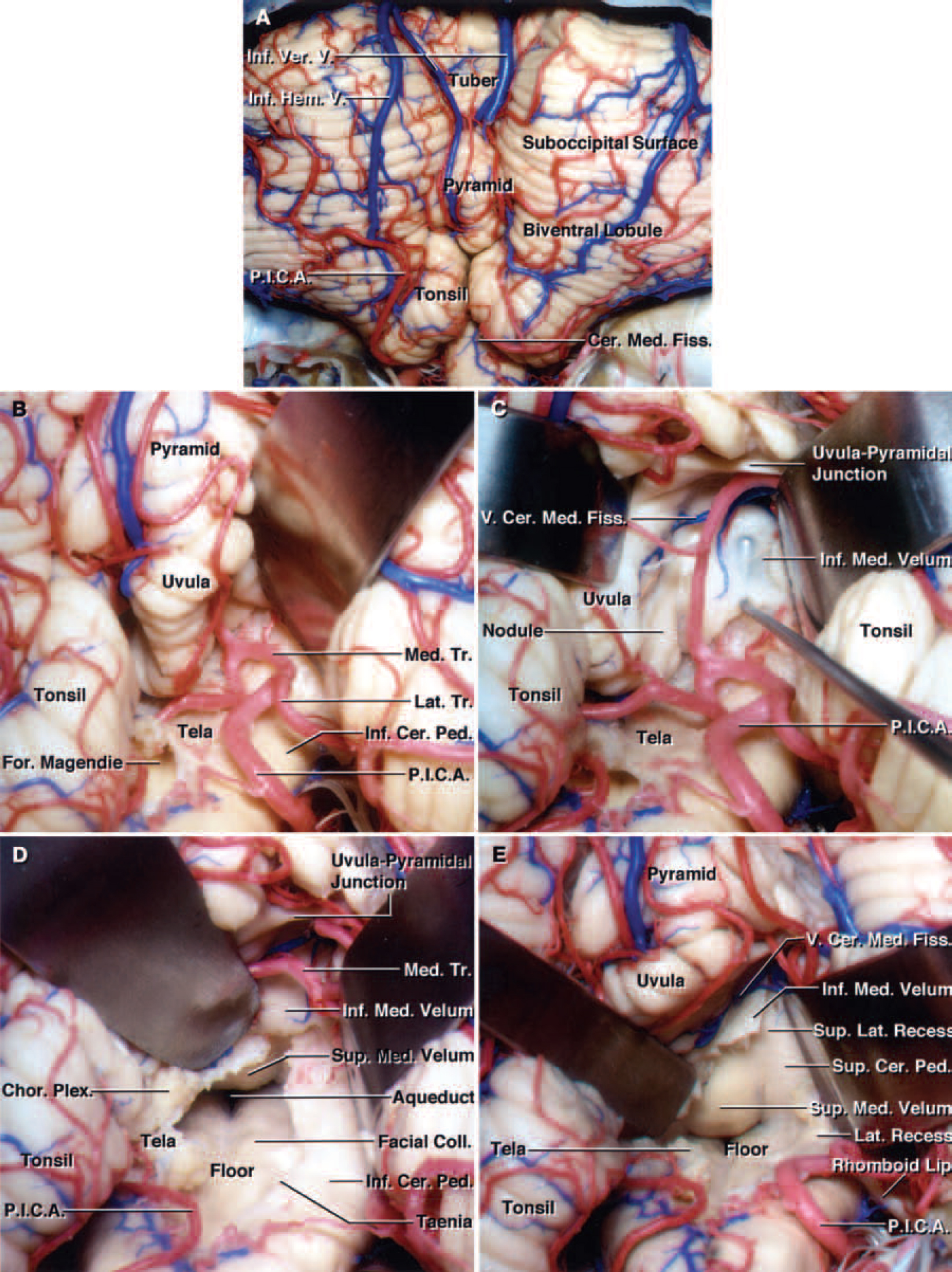
FIG. 6. Photographs of a cadaveric brain specimen showing the telovelar approach to the fourth ventricle. A: After the exposure of the tonsil and biventral lobule, the uvulotonsillar and medullotonsillar spaces on both sides are opened. B: The tonsil has been retracted laterally to expose the right uvulotonsillar space with special attention directed to preservation of the PICA and its bifurcation into the medial and lateral trunks. The PICA bifurcation is located near the inferior end of the uvulotonsillar space, close to the telovelar junction, in aproximately 30% of the specimens. C: The right tonsil has been retracted superolaterally and the uvula medially to expose the right uvulotonsillar space. The tip of a right-angled nerve hook has been placed beneath to the inferior medullary velum in the superolateral recess. The tip of the hook can be seen through the thin inferior medullary velum. D: The uvula has been retracted superomedially and the right tonsil superolaterally to achieve maximum visualization. The PICA bifurcation has been retracted laterally without sacrificing any of the branches of the vessel. If the PICA bifurcation is present at the uvulotonsillar space along the site of the telovelar incision, it is easier to retract it laterally before creating an incision along the tela than to retract it medially in most hemispheres. The incision along the tela choroidea in the right half of the roof has been extended to the telovelar junction. The entire floor of the fourth ventricle from the obex to the aqueduct has been exposed. E: The right half of the inferior medullary velum has been divided to expose the superolateral recess and maximize the exposure along the medial superior half of the roof and superior cerebellar peduncle. Opening the inferior medullary velum increases the exposure of the lower portion of the superior medullary velum near the fastigium. The vein of the cerebellomedullary fissure, which crosses the upper end of the inferior medullary velum, has been preserved. Retracting the right tonsil superolaterally provides access to the lateral recess and exposes the rhomboid lip. Hem. = hemispherical; Ver. = vermian.
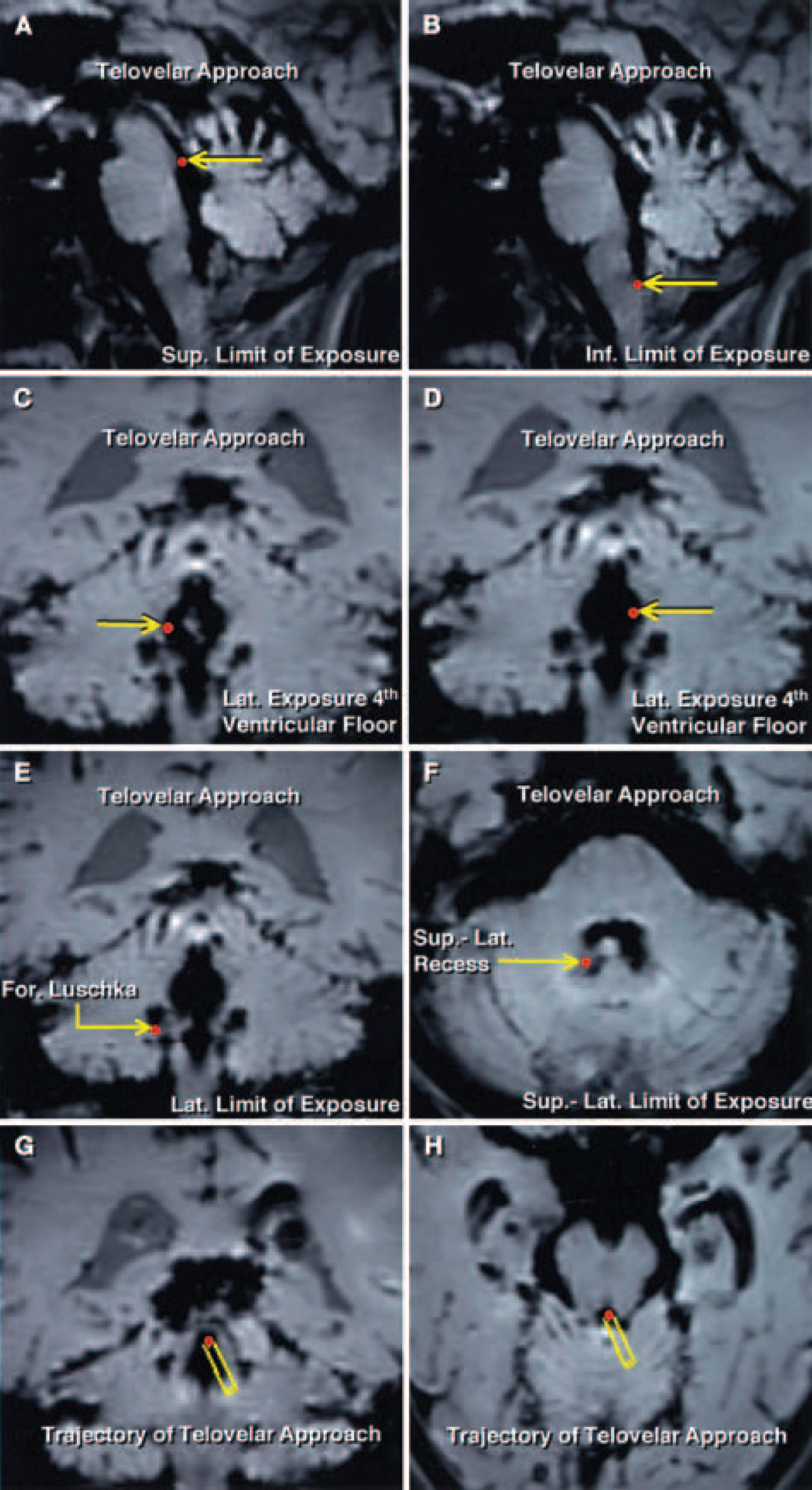
FIG. 7. Telovelar approach. Sagittal, coronal, and axial Stealth MR images of the limits of the surgical exposure (red dots indicated by yellow arrows). A: The superior limit of exposure along the floor of the fourth ventricle is the aqueduct. B: The telovelar approach provides access to the entire length of the floor of the fourth ventricle from the aqueduct to the obex, which is the inferior limit of exposure. C and D: The lateral limits of exposure along the floor of the fourth ventricle are located at the level of the striae medullaris, near the medial edge of the lateral recesses. E: Elevating the tonsils superolaterally provides exposure of the full length of the lateral recess and the foramen of Luschka at the inferolateral end of the lateral recess. F: Superolateral limit of exposure is the superolateral recess, which can be exposed only by incising the inferior medullary velum. G and H: Coronal and axial Stealth MR images obtained during the dissection demonstrating the trajectory of the telovelar approach (yellow rectangles leading to red dots). In the telovelar approach, the fourth ventricle is approached off the midline through the uvulotonsillar space by opening the tela choroidea and the inferior medullary velum lateral to the uvula.

FIG. 8. Photographs of cadaveric brain specimens showing a comparison of the transvermian and telovelar exposures in a stepwise dissection. A and B: Transvermian approach. The transvermian incision directed through the inferior portion of the vermis exposes the underlying nodule, which must be incised to gain access to the fourth ventricle (A). The incision through the vermis was limited by the position of the superior medullary velum, located at the anterosuperior edge of the nodule. Retraction of the paired halves of the lower vermis provides an exposure between the edges of the incised vermis that is approximately 1 to 2 cm wide. Enlarged view (B). Following the opening of the tela choroidea and the inferior medullary velum through a transvermian incision, the full length of the floor (mean length 4 cm) from the aqueduct to the obex, can be exposed. The superior medullary velum limits the upper portion of the exposure. The transvermian approach provides better exposure of the midline portion of the superior half of the roof of the fourth ventricle. The lateral limit of the transvermian exposure extends to the striae medullaris and the medial edge of the vestibular area. C–F: Telovelar approach. The tonsils have been retracted superolaterally to expose the tela choroidea, inferior medullary velum, and both lateral recesses (C). The lateral limit of the exposure achieved is the foramen of Luschka, where the choroid plexus protrudes behind the glossopharyngeal and vagus nerves. The uvula has been retracted medially and the right tonsil laterally, and the tela and inferior medullary velum have been opened to expose the entire length of the floor of the ventricle (D). The aqueduct and the obex are the superior and inferior limits of the exposure along the floor. View of structures shown in panel D (E). The lateral limits of the exposure along the floor of the fourth ventricle are similar to those provided by the transvermian approach and located at the level of the striae medullaris, near the medial edge of the lateral recesses. Incising the inferior medullary velum provides access to the superolateral recess and most of the midline superior half of the roof of the fourth ventricle formed by the the superior medullary velum. View of another specimen (F). Both tonsils have been elevated to expose the full length of the lateral recesses. The rhomboid lip at the lateral end of the lateral recess has been exposed on the right side. The left PICA bifurcates into medial and lateral trunks at the inferior end of the uvulotonsillar space.

FIG. 9. Diagrams superimposed over photographs of a section of cadaveric brain, demonstrating the comparison of operative angles of the transvermian and telovelar approaches along the floor of the fourth ventricle. The angles were measured along the floor of the fourth ventricle in the sagittal plane. The apex of the angle was placed at three sites along the floor of the fourth ventricle: the aqueduct, the midpoint (50% point) of the distance measured from the obex to the inferior border of the aqueduct, and the 75% point of that distance. A: A sagittal section demonstrating the vermian parts of the suboccipital surface of the cerebellum. The highlighted portions of the vermis are the declive (orange), folium (violet), tuber (blue), pyramid (yellow), uvula (green), and nodule (light red). The fastigium, the apex of the roof of the fourth ventricle, divides the roof into superior and inferior portions and is located between the midpoint and the 75% point, near the midsagittal level. The upper limit of the incision (dotted line) extends through the uvula, pyramid, tuber, and the lower portion of the folium to the fastigium, but not into the inferior edge of the superior medullary velum. B: The transvermian and telovelar approaches provide excellent exposure of the entire floor of the fourth ventricle from the obex to the aqueduct. The mean operative angles of the transvermian and telovelar approaches measured at the aqueduct were 38 ̊ (Angle A) and 30 ̊ (Angle B), respectively. In the sagittal plane the exposure gained at the aqueduct when the telovelar approach is used is shaded green. The mean difference between the operative angles of the two approaches at the aqueduct did not exceed 8 ̊ in favor of the transvermian approach (shaded light red). In both the transvermian and telovelar approaches, the operative angle of approach was limited by the superior medullary velum in the sagittal plane. The limitation was most significant when exposing the rostral end of the fourth ventricle near the aqueduct. The working angle decreases when moving away from the midline. In the telovelar approach, the fourth ventricle is approached off the midline, which accounts in part for the decrease in operative angle. The uvulopyramidal junction, the broadest strip of vermis within the posterior incisural space, limited the lateral and medial retractions of the tonsil and the uvula at the uvulotonsillar space, respectively. C: The mean operative angles of the transvermian and telovelar approaches measured at the 75% point were 72 ̊ (Angle A) and 67 ̊ (Angle B), respectively. In the sagittal plane, the exposure at the 75% point for the telovelar approach is shaded green. The mean difference in the working angle gained through the transvermian approach at the 75% point is shaded light red. The lower edge of the superior medullary velum limited the working angle at the 75% point in both approaches and the uvulopyramidal junction accounts for the decrease in the operative angle in the telovelar approach. D: The operative angles of the transvermian and telovelar approaches measured at the midpoint along the length of the floor of the fourth ventricle were identical, 84 ̊ (Angle A). At the midpoint, the working angle gained in the sagittal plane is shaded blue. At the midpoint along the floor of the fourth ventricle, the working angle increased dramatically compared with the working angle at the aqueduct.
Discussion
Approaching the fourth ventricle presents challenges because of its deep location and relationship to vital structures of the brainstem. The transvermian approach has been the most commonly used means to access lesions of the fourth ventricle.3,11,12 An alternative approach directed through the cerebellomedullary fissure to the tela choroidea and inferior medullary velum provides an additional means to access the fourth ventricle and has been used by an increasing number of neurosurgeons.8,16,17,20,21,28 Although there are a few reports in the literature on the microsurgical anatomy of the telovelar approach and its clinical applications,8,20,28 its comparison with the more traditional transvermian approach has not been studied.
A common misconception regarding the telovelar approach is that approaching lesions extending into the upper half of the fourth ventricle is difficult due to a reduced working angle and limited exposure of the region. The goal of the present study was to compare the transvermian and telovelar approaches with regard to working angles and anatomical structures limiting the exposure.
Since Dandy’s statement3 regarding splitting of the vermis without causing a disturbance in function, the transvermian approach with a vermian split has been used extensively.14,25 Nevertheless, the transvermian approach requires an incision into the cortical and functional areas of the cerebellum. The splitting of the inferior cerebellar vermis may cause caudal vermis syndrome, resulting in an equilibratory disturbance with truncal ataxia, gait disturbance, oscillation of the head and trunk, and nystagmus.6,7 Moreover, because the dentate nucleus is located along the posterolateral margin of the roof of the fourth ventricle adjacent and rostral to the superior pole of the tonsil, this nucleus may be damaged when splitting the cerebellar vermis.28 When the dentate nucleus is damaged, the equilibratory disturbances that are created are more severe than those observed with vermian lesions alone and are often accompanied by intentional tremor during voluntary movement of the extremities. Splitting the inferior portion of the vermis may also play a role in cerebellar mutism, a transient complication following the removal of a cerebellar and fourth ventricle tumor. Although the exact anatomical substrate for cerebellar mutism remains unknown, the inferior portion of the vermis, including the pyramid, uvula, and nodule, has been implicated.2,4,5,21,24
The use of natural clefts in the cerebellomedullary fissure to approach the fourth ventricle began after a detailed description of the microsurgical anatomy of the region had been published.17,19 Yasargil26,27 described a median inferior suboccipital approach along the “tonsilouveal” sulcus to treat fourth ventricle lesions; however, the use of the cerebellomedullary fissure to access the fourth ventricle without splitting the vermis was first reported by Matsushima, et al.16 Subsequently, this method and satisfactory results were reported by different authors.10,28 The senior author (A.L.R.) has been using the telovelar approach in his practice for more than two decades and has observed no clinical deficit resulting from the opening of the tela and velum. We have recently described the means of exposing the fourth ventricle through the telovelar approach.20
The transvermian and telovelar approaches are distinct from each other in several key aspects. These distinguishing features include the direction and route of the approach, the structures retracted and sacrificed, the direction of the retraction, the structures limiting the exposure, and the areas exposed. The location and direction of the growth of a fourth ventricle or a brainstem lesion play a role in the selection of the ideal approach. Lesions located predominantly in the vermis and extending into the fourth ventricle are accessed by following the transvermian approach. Tumors located on the ependymal surface and in the floor of the fourth ventricle often do not invade the cerebellum and can be accessed using the telovelar approach. Tumors of the fourth ventricle not infrequently extend to the lateral recess, which can be best approached by elevating the ipsilateral tonsil and using the telovelar route. The transvermian incision has the advantage of being in the midline, but gentle retraction of the uvula to the opposite side places the telovelar approach very near the midline. The obex, the posteromedian sulcus of the medulla oblongata, and the foramen of Magendie at the lower edge of the telovelar and transvermian exposures provide landmarks valuable in orienting the operative view to the midline.
The transvermian approach provides a slightly greater working angle in the sagittal plane when approaching the rostral one fourth of the floor of the fourth ventricle. In addition, a slightly better visualization of the midline inferior portion of the superior medullary velum and fastigium is obtained through the transvermian approach. Nevertheless, the main disadvantage of the transvermian approach is a limited lateral exposure. The lateral limit of exposure is the initial portion of the vestibular area along the floor of the fourth ventricle in the transvermian approach, and the lateral recesses cannot be accessed unless part of the vermis or the tonsils is removed. Another drawback to the transvermian approach is the potential for complications due to iatrogenic neural injury caused by the vermian incision, removal of the vermis or tonsils, and the retraction of two halves of the split vermis.
Matsushima, et al.,17 recently reported on the methods of dissecting the cerebellomedullary fissure and proposed three different opening procedures for the transcerebellomedullary fissure approach: extensive, lateral wall, and lateral recess types of procedures. Following the complete dissection of uvulotonsillar and medullotonsillar spaces, we were able to obtain maximum exposures by using a similar set of incisions. Our initial incision was located along the tela choroidea, adjacent to the taenia, extending superolaterally to the telovelar junction. The opening of the tela was followed by an incision along the inferior medullary velum and the posterior margin of the wall of the lateral recess. The telovelar approach exposes the full extent of the floor of the fourth ventricle in a manner similar to the transvermian approach. In addition, the superolateral recess and structures along the lateral recess, including the foramen of Luschka, can be accessed without removal of brain or adjustment of retractors.
A limitation of exposure and a decrease in the working angle due to two anatomical structures can be expected at the inferior portion of the superior medullary velum near the fastigium. This limitation is due, in part, to the fact that the superior medullary velum forms a more narrow angle to the fourth ventricle floor in its lateral aspect when compared with the midline. In addition, the uvulopyramidal junction located at the rostral end of the uvulotonsillar space limits exposure. The superior medullary velum and the uvulopyramidal junction limited the working angles at the aqueduct and the 75% point along the floor of the fourth ventricle. The difference between the working angles of the two approaches was less than 8 ̊, and this modest decrease in the working angle of the telovelar approach disappeared at the midpoint of the floor of the fourth ventricle. At the midpoint of the floor, the working angles of the two approaches more than doubled compared with the exposure at the aqueduct.
Compared with the transvermian approach, the telovelar approach provides equivalent exposure of the floor of the fourth ventricle and additional exposure of the superolateral recess and the foramen of Luschka, with only a modest decrease in working angles when approaching the rostral one fourth of the ventricle. Nevertheless, the exposure is achieved without removal of or incision through the functional areas of the cerebellum and may be associated with a lower incidence of mutism and other postoperative neurological sequelae.
Contributors: Necmettin Tanriover, MD, Arthur J. Ulm, MD, Albert L. Rhoton, Jr, MD, and Alexandre Yasuda, MD
This article was originally published here: Tanriover N, Ulm AJ, Rhoton AL, Jr, Yasuda A. Comparison of the transvermian and telovelar approaches to the fourth ventricle. J Neurosurg 2004;101:484–498, doi.org/10.3171/jns.2004.101.3.0484, and is included through an exclusive partnership with the Journal of Neurosurgery and its parent company, the American Association of Neurological Surgeons (AANS). The AANS retains full copyright. The appearance of this material here does not imply open access or free use by any other party.
The Neurosurgical Atlas is honored to maintain the legacy of Albert L. Rhoton, Jr, MD.
References
- Cohen AR (ed): Surgical Disorders of the Fourth Ventricle. Cambridge: Blackwell Science, 1996, pp 147–160
- Dailey AT, McKhann GMII, Berger MS: The pathophysiology of oral pharyngeal apraxia and mutism following posterior fossa tumor resection in children. J Neurosurg 83:467–475, 1995
- Dandy WE: The Brain, in Lewis D(ed): Practice of Surgery. Hagerstown, MD: WF Prior, 1966, pp 452–458
- Dietze DD Jr, Mickle JP: Cerebellar mutism after posterior fossa surgery. Pediatr Neurosurg 16:25–31, 1990
- Frazier CH: Remarks upon the surgical aspects of tumors of the cerebellum. NY State J Med 18:272–280, 332–337, 1905
- Holmes G: The Croonian lectures on the clinical symptoms of cerebellar disease and their interpretation. Lancet 1:1177–1182, 1231–1237, 1922
- Holmes G: The Croonian lectures on the clinical symptoms of cerebellar disease and their interpretation. Lancet 2:59–65, 111–115, 1922
- Jean WC, Abdel Aziz KM, Keller JT, et al: Subtonsillar approach to the foramen of Luschka: an anatomic and clinical study. Neurosurgery 52:860–866, 2003
- Johnston TB: A note on the peduncle of the flocculus and the posterior medullary velum. J Anat 68:471–479, 1934
- Kellogg JX, Piatt JH Jr: Resection of fourth ventricle tumors without splitting the vermis: the cerebellomedullary fissure approach. Pediatr Neurosurg 27:28–33, 1997
- Kempe LG: Operative Neurosurgery. New York: Springer-Verlag, Vol 2, 1970, pp 1–13
- Kempe LG: Operative Neurosurgery. New York: Springer-Verlag, Vol 2, 1970, pp 14–33
- Koos WT, Spetzler RF, Lang J: Color Atlas of Microneurosurgery: Microanatomy, Approaches, and Techniques. New York: Thieme Medical, 1993, pp 483–487
- Kyoshima K, Kobayashi S, Gibo H, et al: A study of safe entry zones via the floor of the fourth ventricle for brain-stem lesions. A report of three cases. J Neurosurg 78:987–993, 1993
- Lister JR, Rhoton AL Jr, Matsushima T, et al: Microsurgical anatomy of the posterior inferior cerebellar artery. Neurosurgery 10: 170–199, 1982
- Matsushima T, Fukui M, Inoue T, et al: Microsurgical and magnetic resonance imaging anatomy of the cerebello-medullary fissure and its application during fourth ventricle surgery. Neurosurgery 30:325–330, 1992
- Matsushima T, Inoue T, Inamura T, et al: Transcerebellomedullary fissure approach with special reference to methods of dissecting the fissure. J Neurosurg 94:257–264, 2001
- Matsushima T, Rhoton AL Jr, de Oliveira E, et al: Microsurgical anatomy of the veins of the posterior fossa. J Neurosurg 59: 63–105, 1983
- Matsushima T, Rhoton AL Jr, Lenkey C: Microsurgery of the fourth ventricle: Part 1. Microsurgical anatomy. Neurosurgery 11:631–667, 1982
- Mussi ACM, Rhoton AL Jr: Telovelar approach to the fourth ventricle: microsurgical anatomy. J Neurosurg 92:812–823, 2000
- Pollack IF, Polinko P, Albright AL, et al: Mutism and pseudobulbar symptoms after resection of posterior fossa tumors in children: incidence and pathophysiology. Neurosurgery 37:885–893, 1995
- Rhoton AL Jr: Cerebellum and fourth ventricle. Neurosurgery 47 (Suppl 3): S7–S27, 2000
- Sekhar LN: Midline and paramedian posterior fossa approaches to cerebellar and brainstem lesions, in Sekhar LN, de Oliveira E (eds): Cranial Microsurgery: Approaches and Techniques. New York: Thieme, 1999, pp 378–399
- Van Calenbergh F, Van De Laar A, Plets C, et al: Transient cerebellar mutism after posterior fossa surgery in children. Neurosurgery 37:894–898, 1995
- Weil SM, Tew JM Jr: Surgical management of brain stem vascular malformations. Acta Neurochir 105:14–23, 1990
- Yasargil MG: Microneurosurgery. New York: Thieme, 1996, Vol 4B
- Yasargil MG: Microneurosurgery. Stuttgart: Georg Thieme Verlag, 1988, Vol 3B
- Ziyal IM, Sekhar LN, Salas E: Subtonsillar-transcerebellomedullary approach to lesions involving the fourth ventricle, the cerebellomedullary fissure and the lateral brainstem. Br J Neurosurg 13: 276–284, 1999
Please login to post a comment.




